Inclusions
The most conspicuous inclusions in amoeboid cells are crystals, glycogen bodies, lipid globules, and endosymbionts.
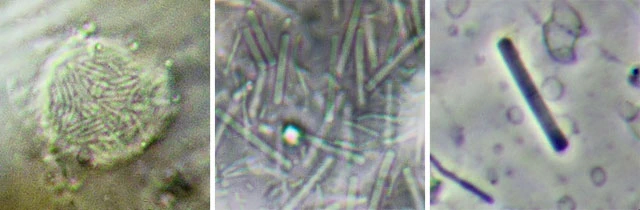
Glycogen bodies
Some large amoebae contain glossy, spheroid glycogen reserves. These structures have been described by various authors as glossy bodies, Glanzkörper, refringent bodies, glycogen bodies, or glycogen spherules. Their size, shape, and number vary among species.
Pelomyxa species, for example, contain numerous refringent glycogen bodies.

Crystals
Crystals are among the most obvious inclusions in many amoebae. Large species such as Amoeba proteus and Polychaos dubium are often filled with numerous crystals, each enclosed within a vacuole. Most freshwater amoebae contain bipyramidal crystals of orthorhombic form, composed of triuret, a nitrogenous waste product. Crystals are produced only by freshwater amoebae; they are absent in Pelomyxa and some other taxa.
Crystal morphology is usually characteristic for a given species, although culture conditions and prey type can influence their appearance.
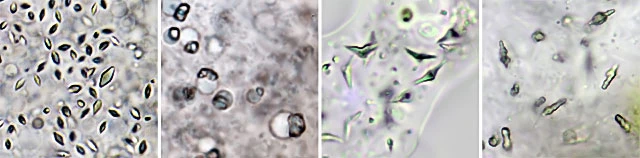

Endosymbionts
Some amoebae contain bacteria or small green algae within their cytoplasm. These organisms form symbiotic relationships with their hosts and are known as endosymbionts. Many large Difflugia species, for example, harbor spherical green algae known as zoochlorellae, sometimes filling the entire cytoplasm. These mixotrophic amoebae typically inhabit nutrient‑poor environments such as peat bogs.
Zoochlorellae provide oxygen and essential nutrients to their host and also serve as a form of “starvation insurance”, as the amoeba can digest its symbionts during periods of food scarcity. The exact mechanisms of this relationship remain unclear in some groups,
One alga to rule them all
Green endosymbionts has been found in three of the five super groups of Eukaryotes: Arcellinida, Euglyphida and
Green endosymbionts have been found in three of the five major eukaryotic supergroups: Arcellinida, Euglyphida, and Stramenopiles. All studied symbionts belong to Chlorella sensu stricto, closely related to Paramecium bursaria symbionts, some lichen symbionts, and several free‑living algae.
Symbionts from all investigated testate amoebae were nearly identical and were assigned to a new trebouxiophycean lineage named TACS (Testate Amoeba Chlorella Symbionts). This “one alga fits all mixotrophic testate amoebae” pattern suggests that these photosynthetic symbionts possess pre‑adaptations for endosymbiosis, enabling them to colonize diverse hosts from a free‑living stage (Gomaa et al., 2014).

Important note: Not all spherical green algae inside an amoeba are true endosymbionts; some may simply be undigested prey.
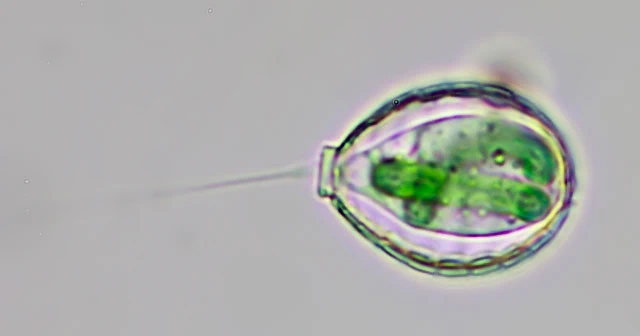
A remarkable exception is the testate amoeba Paulinella chromatophora, which contains a photosynthetic endosymbiont of cyanobacterial origin. This organelle, known as a chromatophore, represents an independent primary endosymbiosis and is now considered a true organelle.
The role of bacterial endosymbionts is not always clear, but the relationship is generally beneficial for both partners. For example, the giant amoeba Pelomyxa lacks mitochondria but contains aerobic bacteria that perform analogous metabolic functions.