Velamentofex dujardini Siemensma and Holzmann, 2023
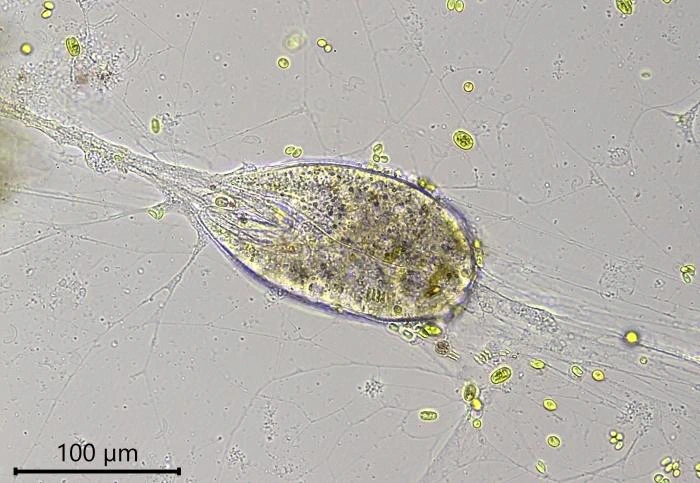
Diagnosis: Multinucleate organic walled monothalamous foraminifera. Shell spherical to pyriform, in some species surrounded by a broad layer of mucus-like material not easily observable as it is covered with food particles, digestion residues and detritus. Shell wall flexible, colorless, and smooth, varying in thickness. Aperture terminal, oblique. Peduncle trumpet-shaped, eccentrically located and varying in length, surrounded by a hyaline peduncular sheath. Radiating network of granuloreticulopodia that can extend several millimeters over the substrate. Nuclei ovular, numerous. Cytoplasm with many contractile vacuoles in the periphery of the cell and numerous refractive rod-shaped yellowish crystalline inclusions, attached to a spherical non-refractive bodies. Cysts observed.
Type locality: River Gardon d’Anduze, Cardet, France (44°01’48.3″N 4°04’27.9″E)
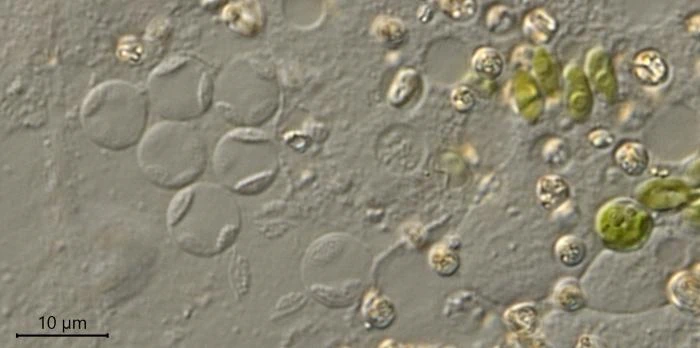
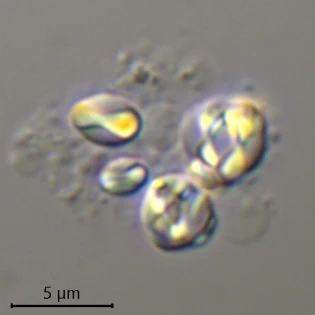
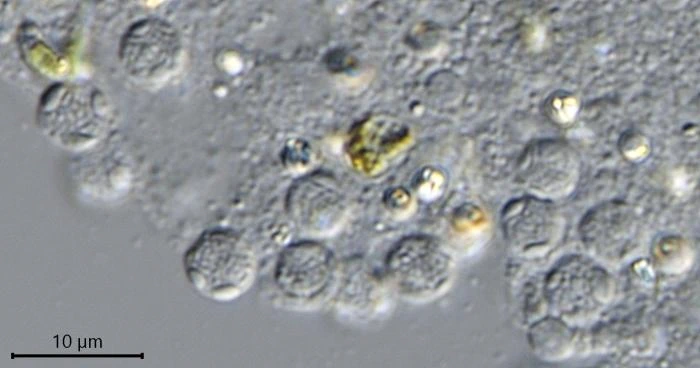
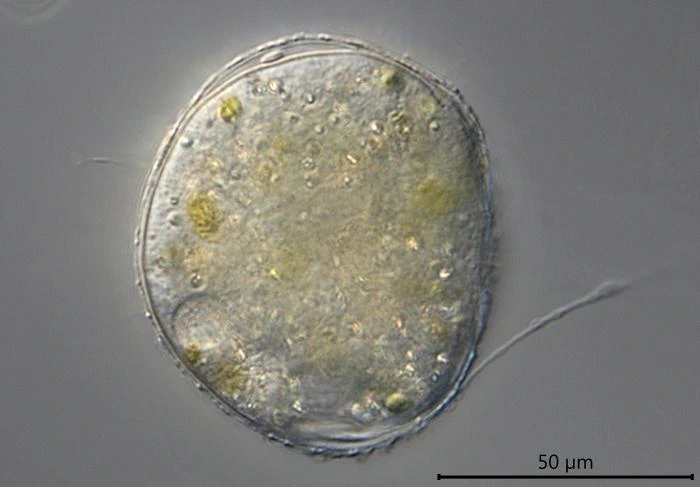
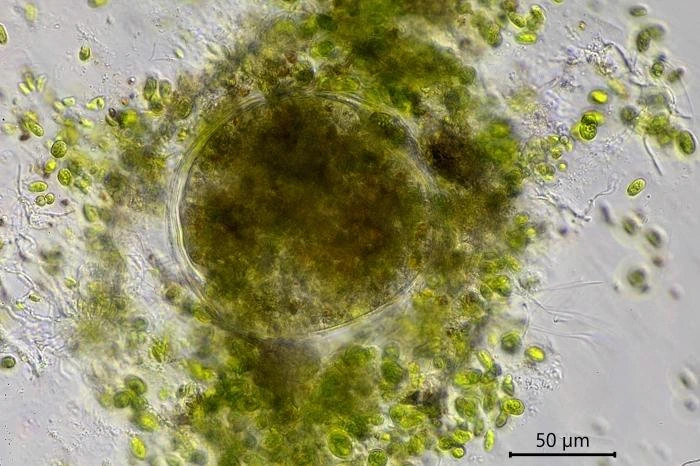
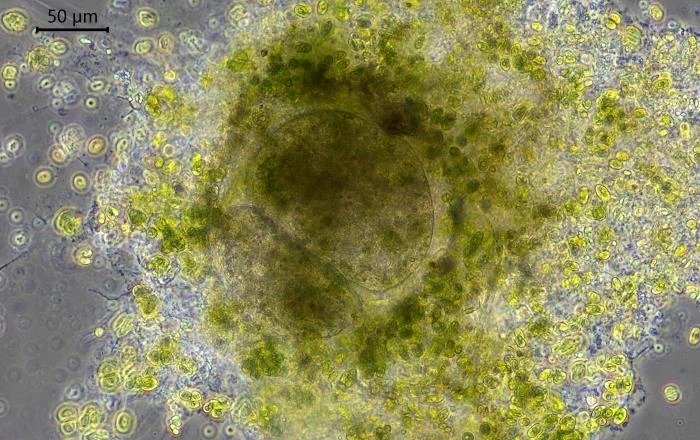
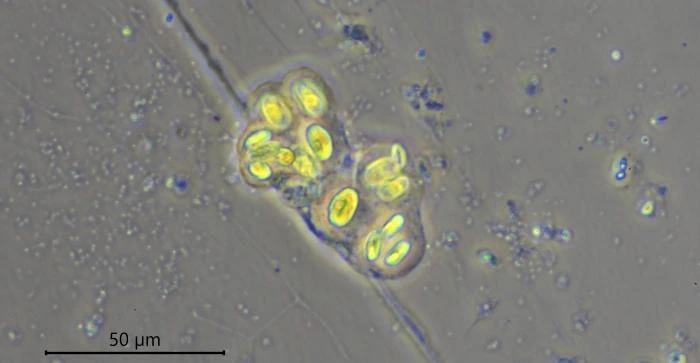
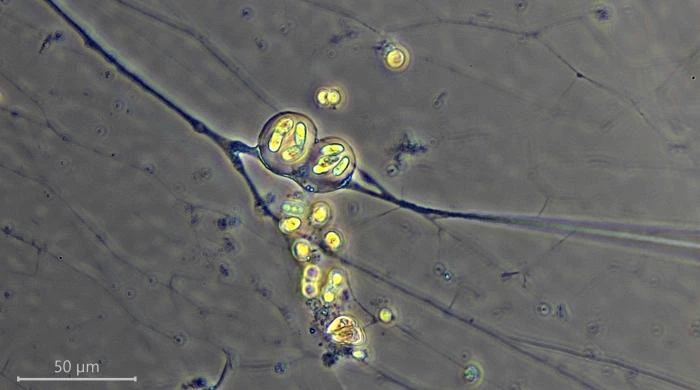
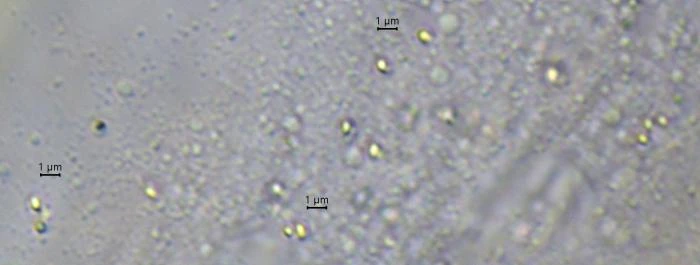
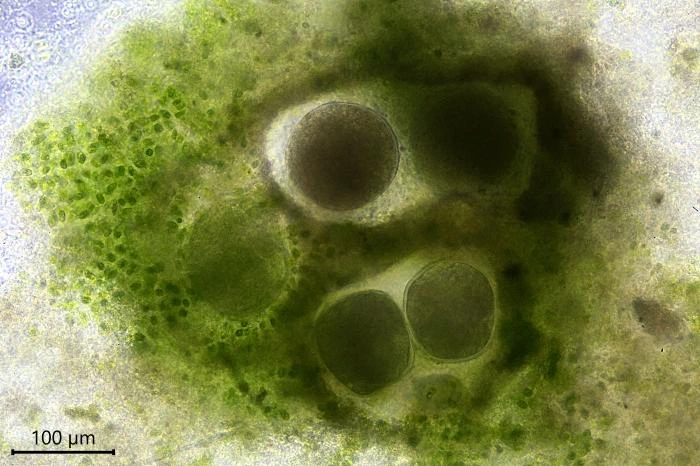
Description: Stationary and sedentary cells are spherical to ovoid, 63-151 µm long and 51-98 µm wide (n=12). Locomotive cells are pyriform to elongate pyriform, ranging in width from 77-87 µm and in length from 83-215 µm (n=12). The cytoplasm contains typical yellow bi-refringent crystalline inclusions attached to spherical bodies. The crystals are rod-shaped and narrowed in the center. Some crystalline inclusions are irregular and two to three can be attached to the same body. Crystalline inclusions and their adhered bodies measure 2.4-2.8 µm in size.
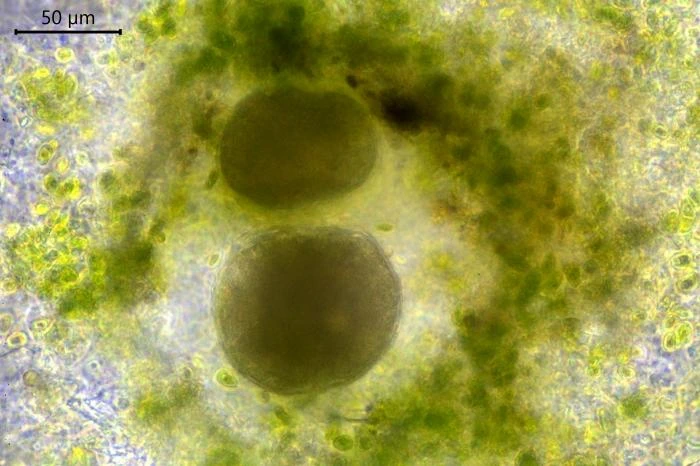
The nuclei are spherical with some lenticular nucleoli, all arranged close to the nuclear membrane. In optical cross-section, 2-3 lenticular nucleoli are visible. It is not clear whether these nucleoli are interconnected, i.e. part of one larger band-shaped nucleolus. One specimen had small spherical peripheral nucleoli, which differed in size. In some specimens, the interior of the nuclei shows a fine granular structure. Nuclei in cysts contain spherical nucleoli scattered throughout the nucleus. The nuclei content in cells ranges from 28 to 150. The diameter of nuclei ranges from 4.6-9.8 µm in trophozoites and from 6.1-7.6 in cysts.
Differential diagnosis. V. dujardini differs from V. berolinensis, V. tyrolensis and V. saxonensis by the absence of a mucus layer. It also differs from these species by the structure of the nucleus, which has a few lenticular nucleoli while the other three species have finely granulated nucleoli (V. berolinensis), some scattered small nucleoli (V. tyrolensis) or a central nucleolus with some nucleoli close to the nuclear membrane (V. saxonensis) (Siemensma et al., 2021). The nuclei in V. berolinensis are much larger, ranging from 11.6−16.2 µm in diameter, versus 4.6-9.8 µm in V. dujardini. The newly described species branches in a clade containing V. berolinensis, V. saxonensis and V. tyrolensis. All species are supported by high bootstrap support (96%-100%), the family Velamentofexidae is equally sustained by significant BV (95%). The obtained sequences of V. dujardini contain 423 to 1168 nucleotides due to different reverse primers used for amplification. The GC content ranges from 33% to 34%.
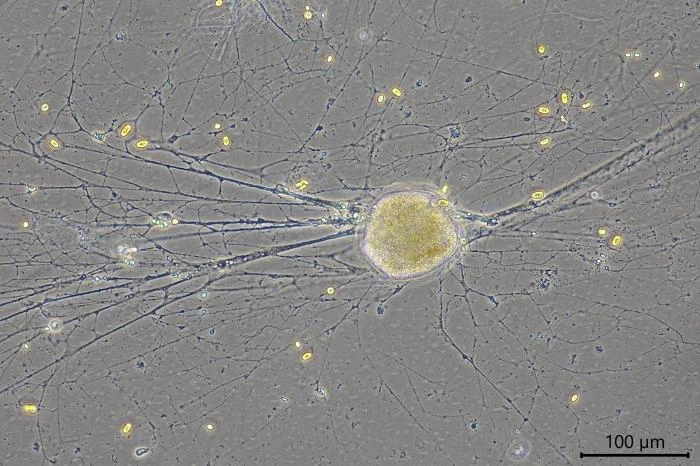
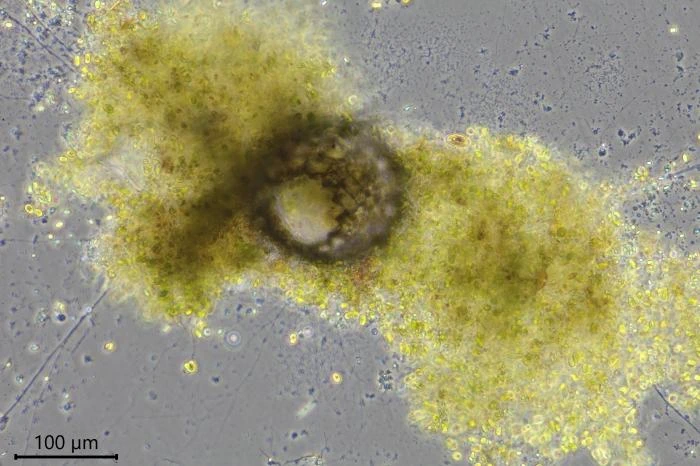
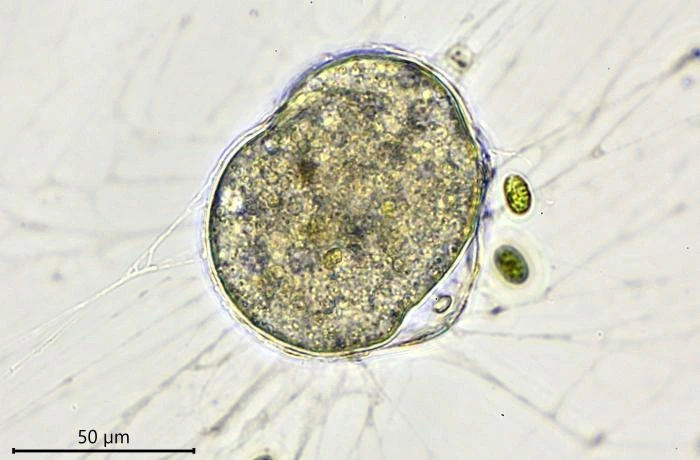
Habitat: Locomotive cells migrate along one thick long granoreticulopodium with one or two long trailing strands. Such a pseudopodial strand can reach a length of more than 1,000 µm. When locomotive cells become stationary, they spread granuloreticulopodia in all directions in search of food. The granuloreticulopodial network of stationary cells forms two to four main strands, sometimes extending more than a centimeter. These main strands regularly change position and vary in length. Within a few days, sedentary cells envelope themselves with food particles and digestive debris, building a protective cover. Cells will also settle in empty testate amoeba shells, e.g. of Netzelia corona, Pseudonebela africana or Difflugia sp., in dead algal cells or empty carapaces of Copepods or Daphnia. Four V. dujardini specimens were observed in one N. corona shell, with one of the specimens departing. The next day, a second cell left the shell. Cells often stay in one place for days or even weeks, sometimes up to three weeks, dividing in their shelter.
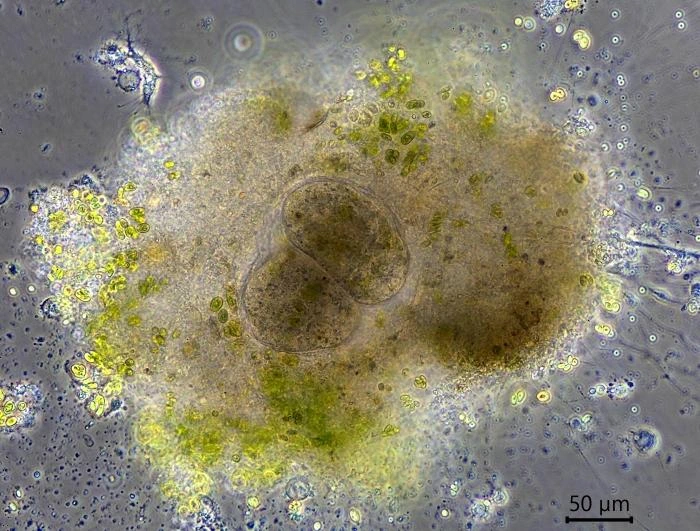
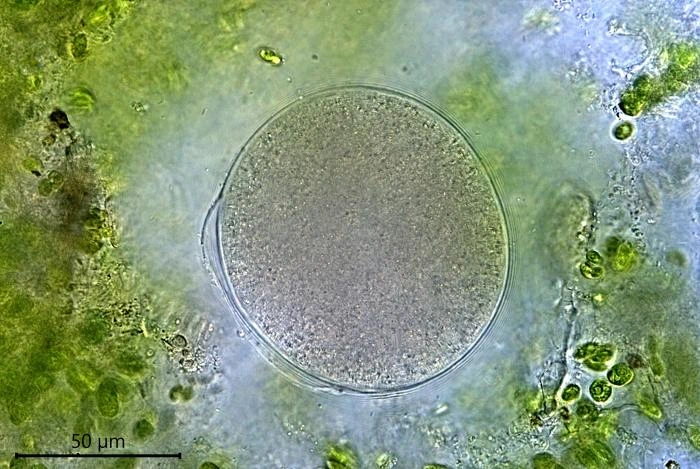
Reproduction. When a cell multiplies by binary division, one of the resulting cells leaves its shelter and moves to a new location, some millimeters or centimeters away. Leaving cells contain no food particles, while the remaining cells are filled with numerous food particles. Sometimes cells remain in the shelter after division and up to five cells can be observed together. When conditions in the petri dish become less favorable, the cells encyst.
Feeding behavior. When starting the culture, only Chlorogonium was added as nutrient and V. dujardini fed on these algae. The algae were also used to build a protective shelter even when cells were already hiding in an empty Difflugia test. After a week, some boiled baker’s yeast was added to the medium and all Velamentofex cells switched to this new nutrient. Chlorogonium cells were then ingested only sporadically. A week later, when small amounts of crusted Tetramin© micro-flakes were added, the Velamentofex cells switched to this new food source. Over the next few days, two naked amoeba species developed in the petri dishes. They were probably introduced as cysts with Tetramin© flakes. One naked amoeba was a vannellid species, the other probably an Angulamoeba. Both amoebae multiplied in the culture dishes with vannellids heavily preyed upon by V. dujardini while Angulamoeba cells were not attacked by them. No other food items were taken up by V. dujardini as long as vannellids were present in the cultures. When a vannellid made contact with a granulopodium, it continued its movement, but the granulopodium remained attached and soon more granulopodia adhered to the vannellid, slowing its movement and causing the amoeba to wrinkle. The wrinkled and later globular prey was transported to the aperture of the Velamentofex cell and ingested.
Remarks: All V. dujardini specimens are surrounded by a clear membranous wall that is flexible, thin, and colorless. No trace of a mucus-like layer is present as reported for the three other described species (Siemensma et al., 2021).